




Modèles d’allocation des ressources : théorie et applications en biotechnologie microbienne
Points forts
- Les modèles d’allocation des ressources fournissent un cadre théorique pour guider l’optimisation.
- Différents modèles d’allocation des ressources sont expliqués et leur applicabilité est examinée.
- Les contraintes qui limitent les objectifs d’optimisation dépendent du contexte et des conditions.
- Le concept d’efficacité du protéome est abordé dans le contexte du métabolisme de débordement.
Les cellules sauvages allouent leurs ressources limitées à l’expression de protéines essentielles à leur croissance et à leur survie dans leur environnement naturel. À l’inverse, la biotechnologie vise à maximiser des indicateurs clés de performance tels que le rendement, la productivité ou le titre. Cependant, cette maximisation se heurte inévitablement à des contraintes physiques, biochimiques, génétiques et évolutives, engendrant des compromis entre des objectifs concurrents. Un défi majeur de la biotechnologie microbienne consiste donc à aligner le comportement cellulaire sur les objectifs de production, ce qui peut être réalisé en manipulant les conditions de culture et les stratégies d’allocation des ressources intracellulaires grâce à une ingénierie métabolique ciblée ou à l’évolution adaptative en laboratoire. Les modèles d’allocation des ressources fournissent un cadre théorique permettant de comprendre et d’orienter ces efforts d’optimisation. Dans cet article, nous passons en revue l’état actuel de la modélisation de l’allocation des ressources, incluant les outils, les méthodes et les fondements théoriques, et nous discutons de leurs applications actuelles en biotechnologie microbienne.
Résumé graphique
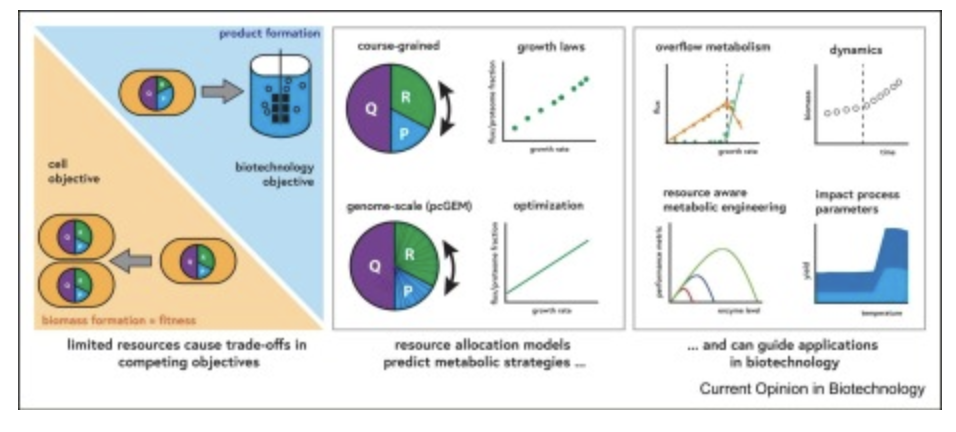
Pourquoi s’intéresser à l’allocation des ressources en biotechnologie ?
La plupart des cellules adaptent leur composition cellulaire et l’expression de leurs protéines en fonction de leur environnement. L’accumulation de données et de théories a permis d’établir un cadre d’allocation (optimale) des ressources pour expliquer et prédire la réussite des adaptations cellulaires, en tenant compte des coûts et des bénéfices liés à l’amélioration de la fitness, qui, chez les micro-organismes, correspond au taux de croissance moyen <sup> 1,2 </sup> . Cependant, pour les applications biotechnologiques, les indicateurs de performance pertinents concernent généralement le rendement, la productivité et le titre d’un produit, ce qui détourne les ressources de la formation et de la croissance de la biomasse. Par conséquent, les objectifs évolutifs de la cellule ne correspondent pas nécessairement à ceux de l’application, ce qui engendre des compromis et peut conduire à une perte de fitness, une stabilité réduite des souches modifiées, des effets pléiotropiques ou une diminution du rendement du produit par rapport à la biomasse.On peut distinguer différents types de coûts liés aux ressources : la charge protéique est liée à l’expression des protéines, tandis que la charge métabolique est liée à la consommation de ressources métaboliques telles que les précurseurs de carbone ou l’ATP. Un exemple de charge protéique, connu depuis les années 1950, est l’expression de protéines hétérologues qui réduit directement le taux de croissance et peut entraîner un métabolisme de débordement [3] , un phénomène qui, curieusement, est généralement observé à des taux de croissance élevés. Étant donné que, chez les bactéries, la densité protéique est souvent constante quelles que soient les conditions [4] , l’expression de protéines hétérologues remplace d’autres protéines sans coût métabolique supplémentaire, sauf si la composition en acides aminés s’écarte de la moyenne. Les protéines sécrétées, en revanche, engendrent une charge pour la machinerie de synthèse protéique, ainsi qu’une charge métabolique, mais n’occupent pas d’espace protéique dans la cellule elle-même. Les produits non cataboliques, c’est-à-dire les produits dont la synthèse nécessite de l’ATP, tels que les molécules plus volumineuses [5] , imposent généralement à la fois une charge protéique due aux enzymes exprimées de manière hétérologue et une consommation supplémentaire d’ATP et de métabolites.Aujourd’hui, nous disposons d’une vision beaucoup plus quantitative et précise des compromis résultant d’objectifs concurrents, notamment pour des organismes modèles tels que *E. coli* et *S. cerevisiae* . Ces avancées sont largement dues aux modèles informatiques, grâce à la disponibilité croissante d’outils et de méthodes <sup> 6,7,8 </sup> . L’ encadré 1 présente les principes de base, et le tableau 1 offre une vue d’ensemble des approches actuelles, de leurs hypothèses, de leurs limites et de leurs applications.
Case 1 Modèles et théories de l’allocation des ressources.Dans les modèles d’allocation des ressources, l’allocation du protéome est étudiée dans le cadre d’un budget protéique contraint, avec une fraction protéique donnée indépendante de la croissance et une répartition de la fraction restante entre différents processus cellulaires (modèles à gros grains) ou des protéines individuelles (modèles basés sur pcGEM), voir Figure 1. Les principales différences entre les approches sont (i) les modèles linéaires basés sur des contraintes par rapport aux modèles cinétiques ; et (ii) les modèles basés sur des données/lois de croissance phénoménologiques ou sur l’optimisation.
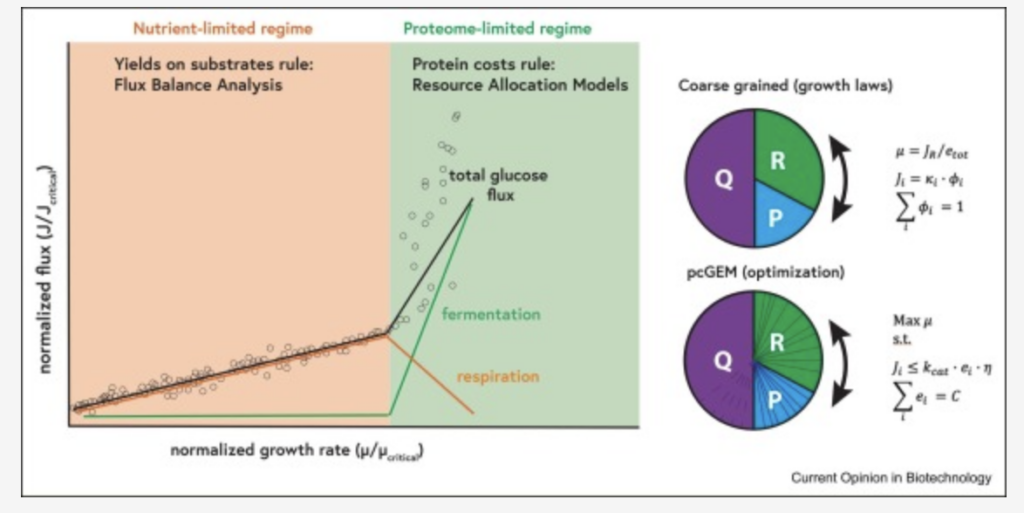
Figure 1. Allocation
du protéome selon deux approches : l’une phénoménologique (basée sur les lois de croissance) (en haut), où les secteurs du protéome sont considérés comme un ensemble, et l’autre à l’échelle du génome (en bas), où chaque protéine est modélisée séparément, mais peut être assignée à un secteur du protéome. Dans les deux cas, le secteur Q (protéines non utilisées) est fixe, tandis que les secteurs R (fraction ribosomique) et P (protéines favorisant la croissance) varient selon les conditions. La formulation mathématique principale est indiquée, avec m le taux de croissance, J <sub>R</sub> le flux ribosomique, k la constante de vitesse phénoménologique, f la fraction du protéome, e la concentration enzymatique et h la saturation de l’enzyme.
| Méthode | Hypothèses | Avantages | Limites | Applications |
|---|---|---|---|---|
| Lois de croissance / modèles phénoménologiques | Flux proportionnel à l’allocation/l’investissement | Des solutions analytiques sont disponibles selon la mise en œuvre ; une logique dynamique/réglementaire peut être intégrée. | À gros grains ; explicatif, non exploratoire | Aperçu fondamental des stratégies réglementaires (connues) |
| Auto-réplicateurs | Optimisation du taux de croissance ; croissance équilibrée | La cinétique étant prise en compte, la dynamique intracellulaire peut également l’être. | À granularité grossière ; explicatif, non exploratoire ; nécessite une optimisation | Aperçu fondamental des stratégies réglementaires (connues) |
| Modèles à contraintes enzymatiques (EC) | Limite supérieure du flux fixée par Vmax = kcat [ e ] ; optimisation du taux de croissance ; croissance équilibrée | À l’échelle du génome ; prédictions améliorées de l’analyse de la fonctionnelle de la densité (FBA) ; contraintes supplémentaires, toutes linéaires (calcul rapide) | Dépend de paramètres cinétiques (incertains) ; nécessite une optimisation | Intégration des données protéomiques ; prédiction des stratégies métaboliques |
| Modèles ME, RBA et PC | Voir le modèle EC, ainsi qu’une prise en compte explicite de la synthèse protéique | Aucune donnée omique requise, mais prédite | Voir le modèle EC ; introduit des relations non linéaires (plus coûteux en calcul) | Stratégie réglementaire complète / analyse coûts-avantages pour une croissance équilibrée |
| pcGEM dynamiques | Voir ME, RBA, PC, mais sans croissance équilibrée | Conditions dynamiques (ex. périodes d’abondance/de disette), régulation spatio-temporelle (ex. cycle cellulaire) | Voir les modèles ME, RBA et PC ; calculs coûteux | Stratégie réglementaire complète / analyse coûts-avantages en contexte dynamique |
Sur la nature des ressources et des contraintes
Les ressources proviennent en définitive des nutriments, et les cellules répondent par l’expression du répertoire protéique qui utilise ces nutriments pour croître et synthétiser des produits. Le taux de croissance étant limité, il existe nécessairement des contraintes, en fin de compte, au niveau de l’expression des protéines, qui sont la cible de l’évolution. D’autres contraintes, telles que la taille cellulaire, les limitations de diffusion et d’autres contraintes thermodynamiques, cinétiques et génétiques, peuvent généralement être reformulées en termes de contraintes sur l’expression des protéines [1] .La question de savoir quelles contraintes sont « actives », c’est-à-dire celles qui limitent l’objectif de la cellule (ou du biotechnologiste), est cruciale et dépend des conditions de croissance spécifiques. Par exemple, dans le cas d’un chémostat à limitation de carbone et à faible taux de dilution, l’expression du système de transport concerné limite la disponibilité du carbone [9] et donc la croissance. Il ne s’agit probablement pas d’une contrainte directe liée, par exemple, à l’espace membranaire, car les expériences d’évolution en laboratoire montrent généralement une régulation positive de ces systèmes de transport. La croissance et la production sont déterminées par la quantité de carbone disponible. En termes de modélisation, il s’agit d’un régime limité par les nutriments [10] , voir Figure 1 (notez que plusieurs nutriments peuvent être limitants [11] ), où des stratégies à haut rendement sont prédites et observées. Dans ce régime de culture en milieu limité en nutriments, généralement obtenu par des cultures en fed-batch à faible concentration de glucose [3] , la surexpression de protéines hétérologues ou de protéines de résistance au stress n’entraîne qu’un faible coût adaptatif, la croissance étant alors limitée par l’absorption du substrat plutôt que par les ressources protéiques cytosoliques. Par conséquent, l’allocation des ressources est principalement régie par des contraintes de bilan massique stœchiométrique, ce qui signifie que l’analyse classique des voies métaboliques stœchiométriques et l’analyse des flux métaboliques (FBA) peuvent guider efficacement l’ingénierie métabolique.En revanche, en culture discontinue où les nutriments sont en excès, les taux de croissance/production sont limités par des contraintes intracellulaires : c’est ce qu’on appelle le régime limité par le protéome [10] . Dans ce cas, l’expression d’une protéine se fait au détriment d’autres protéines. Par conséquent, allouer davantage d’enzymes à la voie métabolique du produit peut certes augmenter le flux, mais réduit également l’espace protéomique d’autres secteurs fonctionnels, ce qui peut entraîner des effets pléiotropiques, une « reprogrammation métabolique » et une réduction du taux de croissance. Dans ce contexte, les modèles d’allocation des ressources (MAR, voir encadré 1 ) permettent d’expliquer des phénomènes que les modèles purement stœchiométriques ne peuvent pas expliquer.L’un des exemples les plus marquants est le métabolisme de débordement, notamment l’effet Crabtree et Warburg (bactérien). Le passage de la respiration à la fermentation, observé chez de nombreux microbes mais aussi dans les cellules de mammifères à réplication rapide (cellules épithéliales, cellules immunitaires activées, cellules cancéreuses), s’explique par une meilleure efficacité protéomique de la fermentation : elle génère un flux d’ATP plus élevé par protéine investie <sup> 9,12,13 </sup> ( voir encadré 2 ). Le métabolisme de débordement étant utilisé pour la production de nombreux composés, notamment le glutamate et d’autres acides aminés par <i>Corynebacterium glutamicum</i>, il serait intéressant d’appliquer les modèles de métabolisme de débordement (RAM) à ces cas.
